

肠道碱性磷酸酶解毒脂多糖并预防炎症在肠道微生物群的响应
来源:
|
作者:詹妮弗·M·贝茨1,Janie Akerlund,Erika Mittge,和Karen Guillemin
|
发布时间: 2020-06-23
|
15212 次浏览
|
🔊 点击朗读正文
❚❚
▶
|
分享到:
脊椎动物的肠道菌群中含有丰富的脂多糖(LPS)或内毒素。我们证明,刷状缘酶-肠碱性磷酸酶(Iap)在斑马鱼肠道菌群形成过程中被诱导从而在促进黏膜对肠道细菌的耐受中起着关键作用。我们证明,iap缺乏的动物对lps的毒性反应是通过Myd88和肿瘤坏死因子受体(Tnfr)介导的机制来感应的。我们进一步证明,内源性微生物群通过一个涉及myd88和tnfr的过程建立了肠道中性粒细胞的正常稳态水平。iap缺乏的动物表现出过多的肠道中性粒细胞流入,类似暴露于lps的野生型动物。但在无细菌饲养的情况下,缺乏Iap的动物的肠道内缺乏中性粒细胞,这表明iap具有防止肠道细菌炎症反应的功能。
and immunity 2005;73:4309–4314. [PubMed: 15972524]
Wallace KN, Akhter S, Smith EM, Lorent K, Pack M. Intestinal growth and differentiation in zebrafish. Mech Dev 2005;122:157–173. [PubMed: 15652704]
Waymire KG, Mahuren JD, Jaje JM, Guilarte TR, Coburn SP, MacGregor GR. Mice lacking tissue nonspecific alkaline phosphatase die from seizures due to defective metabolism of vitamin B-6. Nature genetics 1995;11:45–51. [PubMed: 7550313]
Westerfield, M. The zebrafish book a guide for the laboratory use of zebrafish Danio (Brachydanio) rerio. Eugene, OR: Institute of Neuroscience University of Oregon; 1993.
Xuan D, Nicolau DP, Nightingale CH, Quintiliani R. Circulating tumor necrosis factor-alpha production during the progression of rat endotoxic sepsis. Chemotherapy 2001;47:194–202. [PubMed: 11306788]
Youngner JS. Bacterial lipopolysaccharide: oral route for interferon production in mice. Infection and immunity 1972;6:646–647. [PubMed: 4628903]
Zambonino Infante JL, Cahu CL. Ontogeny of the gastrointestinal tract of marine fish larvae. Comp
Biochem Physiol C Toxicol Pharmacol 2001;130:477–487. [PubMed: 11738635]
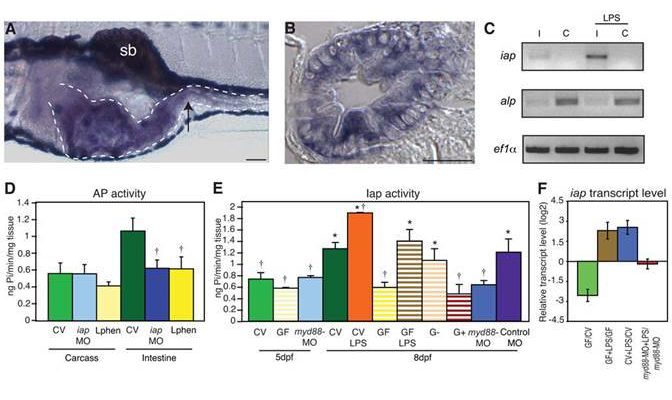
Figure 1.
Iap activity and iap transcription are regulated by LPS. In situ hybridization of iap transcript at 5 days post fertilization (dpf) in (A) a whole mount larva, and (B) a transverse section through the mid intestine (at the point indicated by the arrow in A). The iap specific purple stain is present in the intestinal epithelium (outlined with the dotted line in A) and is distinct from the black pigment cells above and below the digestive tract and the swim bladder (sb) in A. Scale bar in panel A = 100 μm, scale bar in panel B = 5 μm. (C) Semi-quantitative RT-PCR analysis showing iap and alp expression in dissected intestines (I) and carcasses from which intestines were removed (C) of 8 dpf larvae untreated or exposed to 50 μg/ml LPS for 24h. Levels of the housekeeping gene ef1α are shown as an amplification and loading control. (D) AP activity in the carcasses and intestines of 8 dpf untreated CV WT larvae, iap-MO injected larvae or larvae exposure to 10mM L-phen from 5 dpf. † Indicates values that differ significantly as compared to the CV levels of each group (carcass or intestine), P<0.01. (E) AP activity in 5 and 8 dpf intestines from larvae reared CV (solid bars) left untreated, exposed at 5 dpf to 30 μg/ml LPS, or injected at the 1 cell stage with myd88-MO or the control galT-MO, or larvae reared GF (striped bars) left untreated, exposed at 5 dpf to 3 μg/ml LPS, or mono-associated at 5 dpf with a Gram-negative Aeromonas species (G−) or a Gram-positive Streptococcus species (G+). † Indicates values that differ significantly from CV at 8dpf, * indicates values that differ significantly from GF at 8 dpf, P<0.01. For D and E, n=10 dissected intestines/treatment for each trial, with at least 2 trials per treatment. Error bars represent standard deviation. (F) iap transcript levels, measured by qRTPCR, were reduced in 8 dpf GF versus CV animals, and elevated in 8 dpf CV and GF animals exposed for 24h to 30 μg/ml LPS, but not in 8 dpf myd88-MO injected animals reared CV and exposed for 24h to 50 μg/ml LPS. Data are representative of two repeated trials, in which all samples were run in triplicate. Error bars indicate standard deviation. All animals were WT unless otherwise indicated.
下一篇:
无
